












基本信息
- 项目名称:
- 对郎飞结传导节能的怀疑及其解释
- 来源:
- 第十二届“挑战杯”省赛作品
- 小类:
- 生命科学
- 大类:
- 自然科学类学术论文
- 简介:
- 本研究首先以郎飞结的最新科学研究结构为出发点,用物理模型的方式来比拟郎飞结之间的传导过程。然后通过物理模型中各种物理量的变化和实际郎飞结所需物理量的变化的对照和分析。随后大胆总结和概述出“对郎飞结传导节能怀疑”这一结论。最后启发式地由物理模型的研究形式转化为对实际郎飞结传导的实验探索的追问。
- 详细介绍:
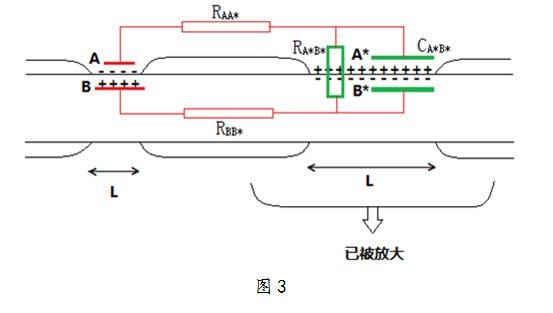
- 一.经典理论认为有髓神经纤维节能传导的原因是髓鞘在有髓神经上占据轴突膜上的Na+泵、K+泵的位置而减少了Na+、K+离子流动的数目,从而减少通过耗能Na-K泵的Na+、K+离子数目,继而减少了所耗ATP的数目,实现了郎飞结处的传导节能。但是由于物理传导模型的推理和郎飞结处高密度Na+泵通道的研究结果,所以对郎飞结传导节能产生了怀疑。 二.郎飞结间的传导及其物理模型 (1)动作电位的产生 由于构成髓鞘的脂质是不导电或不允许带电离子通过的,只能在髓鞘暂时中断的郎飞结处轴突膜才能和细胞外液接触,使跨膜离子移动得以进行[4]。因此有髓纤维受到外来刺激时,只有郎飞结处发生动作电位,从而增加了内向Na+的通透性,使Na+在瞬间不消耗能量的情况下流入轴突膜内,造成膜内正外负的效果。若刺激为阈刺激,就会产生动作电位,膜内外存在动作电压,膜同侧与其有髓神经纤维的其它部位也存在电压。 (2)相邻郎飞结之间的传导及物理传导模型的建立 脂质双层膜存在电阻和电容。膜电阻取决于它们运载粒子流的能力,当存在驱动力的时候—正如铜线的电阻取决于它运载电子电流的能力一样。细胞膜两侧的电位差,使离子通过通道由一侧转移到生物膜的另一侧,通道密度越高,其电阻越低,而电导越高。 膜电容介质来自脂质双层疏水性的内层,这内层是不良导体。当电荷被一个不导电的膜层电容器分隔时,正电荷和负电荷附着并堆积在脂质双层的双侧。分隔的电荷跨脂质双层产生电位差,电位差的大小取决于它的性质。因为产生电压的电荷是按一定的速率提供的,所以膜电容影响跨脂质双层电压变化的时程。 在电学上一片膜可用平行的RC(如图1-A,B)电路代表:电阻和电容平行[5]。图示中,电阻为各个通道引起的电阻,是在正常生理状态下的细胞内部与外部交流的通道,其中不包括K+通道和引起动作电位的Na+通道等类似通道,因为正常静息电压不能使其激活;电容处也存在通道,即电压门控的Na+通道和K+通道等类似通道,只有电压达到一定的电压值时才可以激活开放。 A B 图1 由以上分析,当郎飞结处受到阈刺激爆发动作电位而使膜内外存在电压时,如图2膜外电势为ψA,膜内电势为ψB。在正常生理状态下,相邻郎飞结处膜外的电势为ψA*,膜内的电势为ψB*。显然A、A*之间存在电势差,B、B*之间也存在电势差。由于膜两侧都是导电的,且|ψA‐ψB|〉|ψA*‐ψB*|,所以A*、B*之间存在相对平衡的离子流动,从而建立了一个郎飞结间传导的物理模型,如图3. 图2 图3(见附图) 通路的建立使A-B为A*-B*增大了逆的电化学梯度,A*-B*间离子移动逐渐降低,逐渐使A*-B*发生逆的电位。RAA*、RBB*、RA*B*分担电压消耗电能。A-B间电能逐渐减少。UAB由Uab1减小为Uab2(AB动作电压为Uab1)。 A-B通过轴突能导电的液体给A*B*电容充电,最后充电电压为Uab2,显然Uab1>Uab2。 因为神经细胞中Na+、K+离子通道为电压门控通道[3],所以若A*-B*间产生动作电位,首先须UA*B*大于或等于可引起电压门控通道开放的最小电压值Umin,所以Uab2≥Umin,Uab1>Umin。 假设Uab2≥Umin,A*、B*轴突膜处电压门控通道开放使Na+进入膜内,K+流入膜外,爆发动作电位,则爆发动作电位后的A*-B*即便成为类似A-B的电池,从而为下一次郎飞结处电压门控通道提供所需电压,以此循环下去传导刺激。 由有髓神经纤维郎飞结处传导模型的建立联系无髓神经纤维处传导。因为无髓神经纤维没有髓鞘使其表面相断,所以无髓神经纤维轴突膜处AB与A*B*相距较近,无RAA*和RBB*,且RA*B*与电容CA*B*并联。电池AB给电容CA*B*充电时几乎无能量损失,UAB=Umin。 (3)动作电位产生到恢复正常生理状态期间离子转运及能量消耗 Na+内流去极化爆发动作电位,K+外流复极化使膜内外侧恢复电荷于膜内外相对平衡的生理状态,但是此时的生理状态相比刺激前的正常生理状态,膜内Na+增多,K+减少。于是,在随后由于Na-K泵的作用使Na+、K+数目恢复到刺激前的正常生理状态数目。值得注意的是,Na-K泵每运输3Na+和1K+便消耗1ATP[3],此过程是耗能的。 三、对郎飞结传导节能的怀疑及其分析 经典理论认为有髓神经纤维节能传导的原因是髓鞘在有髓神经上占据轴突膜上的Na+泵、K+泵的位置而减少了Na+、K+离子流动的数目,从而减少通过耗能Na-K泵的Na+、K+离子数目,继而减少了所耗ATP的数目,实现了郎飞结处的传导节能。但是由于以上物理传导模型的推理和郎飞结处高密度Na+泵通道[6]的研究结果,所以对郎飞结传导节能产生了怀疑。 在无髓神经,因为AB与A*B*相距较近使无髓神经爆发产生动作电位的电压最小是Umin,而在有髓神经中,由于RAA*、RBB*使产生的动作电位的电压值将大于Umin,,从而得出若在有髓神经中,刺激所产生的传导效果被传导,则产生动作电位处的郎飞结必须有更多的Na+流入膜内。且有关研究表明郎飞结处高密度Na+通道[6],从而更加有力证明郎飞结处产生的动作电压值大于同等长度无髓神经轴突处产生的电压值,也就是说产生的动作电压越大,内流Na+的数目越多。 因而对于郎飞结不能判断性地认为髓鞘在有髓神经纤维上占据了轴突膜上Na+泵、K+泵、Na-K泵的位置而使其传导产生节能的效果。而应该从总体上比较刺激在同等长度的有髓神经和无髓神经上传导造成的钠离子内流数目和Na-K泵消耗ATP的数目的多少。
作品图片


作品专业信息
撰写目的和基本思路
- 本研究目的是对郎飞结传导节能的怀疑及其解释。 本研究首先以郎飞结的最新科学研究结构为出发点,用物理模型的方式来比拟郎飞结之间的传导过程。然后通过物理模型中各种物理量的变化和实际郎飞结所需物理量的变化的对照和分析。随后大胆总结和概述出“对郎飞结传导节能怀疑”这一结论。最后启发式地由物理模型的研究形式转化为对实际郎飞结传导的实验探索的追问。
科学性、先进性及独特之处
- 作品在对郎飞结传导的节能怀疑的论述中,充分利用所建立的物理模型电传导机制来描述郎飞结间的传导过程,并用物理模型的理论对怀疑做了论证。通过怀疑和物理模型的论述,可使人们对郎飞结的传导有一个全新的认识。 利用物理模型来研究问题较比实验研究有了创新性和简便性,同时结合实验研究结果的科学性而使物理模型的研究有了说服性,也为以后科学研究开辟了一条新的道路——物理模型的研究方法。
应用价值和现实意义
- 近现代研究表明老年性痴呆病、格林-巴利综合征和多发性硬化病与郎飞结的病变有关,也许对郎飞结处电传导和结构的清楚研究和认识会对这些病的治愈具有积极的启发作用。 为以后科学研究开辟了一条新的道路——物理模型的研究方法。
学术论文摘要
- 经典理论认为有髓神经纤维节能传导的原因是髓鞘在有髓神经上占据轴突膜上的Na+泵、K+泵的位置而减少了Na+、K+离子流动的数目,从而减少通过耗能Na-K泵的Na+、K+离子数目,继而减少了所耗ATP的数目,实现了郎飞结处的传导节能。但是由于物理传导模型的推理和郎飞结处高密度Na+泵通道的研究结果,所以对郎飞结传导节能产生了怀疑。
获奖情况
- 暂无,正在准备投稿中。
鉴定结果
- 有待更多的科学理论和实验进行验证
参考文献
- [1]李继硕主编,神经科学基础[M],北京:高等教育出版社, 2002,186-187 [2]江南,齐建国,郎飞结及其周围结构分子组成及其相互作用,《神经解剖学杂志》,2010,26(4);451-454 [3]岳利民,崔慧先主编,人体解剖生理学,第5版,人民卫生出版社,2007,39-49 [4]汪仁斌,刘兴洲,神经传导阻滞及其发生机制[M].脑与神经疾病杂志,2006,14(3):239-241 [5]陈宜张主编,分子神经生物学,人民军医出版社,1995,85-86 [6]Peles E,Salzer JL.Molecular domains of myelinated axous[J],CurrOpin Neurbiol,2000,10(5):5582565 [7]Ratcliffe CF,Westenbrock RE,Curtis R,et al. Sodium channel1β1 and β3 subanits associate with nearofascin through their extracellular immunoglobulin like domain [J],Cell Biol,2001,15(4):4272434
同类课题研究水平概述
- 以往人们主要关注和研究郎飞结在神经冲动传导过程中的作用,例如参与著名的“跳跃式”神经冲动传导机制,而近几年来,随着分子生物学研究的兴起和深入,郎飞结的形成、发育、分子组成、轴突胶质相互作用以及其他方面的功能,引起了人们的广泛兴趣和关注。但是,没有大多资料和课题对郎飞结进行返璞归真的研究,即郎飞结的传导节能等问题的研究。而且现今用物理模型的方法来处理问题的研究少之又少。







